Christoph Nothdurft - Commented Bibliography
This overview lists my major publications, not always in chronological order. (See publications.html for a complete and chronologic list.) Papers are commented and their main findings summarized. Altogether, the list may thus give an overview of the lines and goals of research I had been following during the last 40 years.
Note: Only very few papers are open access and can be downloaded directly. But of most other papers there exist PDFs which I would be happy to send out quickly upon request; please contact me.
1. PHYSIOLOGY OF CORTICAL AND PRECORTICAL CELLS:
EXPLORATION OF SPATIAL PROPERTIES, COLOR SENSITIVITIES, AND TEXTURE REPRESENTATION
•
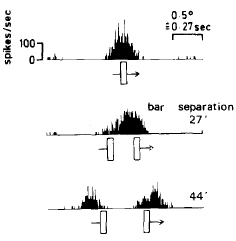
Nothdurft, H.C. (1976).
Spatial resolution and nonlinearities of simple cells in the cat visual cortex measured with parallel line pairs.
Biological Cybernetics, 24, 153-163.
Studied spatial resolution (minimum separabile) of cortical cells. Two kinds of non-linear interactions
were observed when lines are close together: (a) the (partial) suppression of one or the other line; (b) an
apparent pulling together and pushing apart of lines and line responses (in principle corresponding to an angle enhancement like in
the Poggendorf and Zöllner illusions). Both effects were simulated in various models; only a presumed (cortical) interaction
model could explain all non-linearities seen.
•
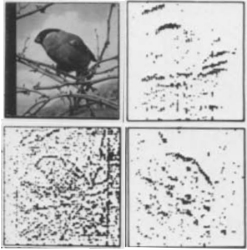
Creutzfeldt, O.D., and Nothdurft, H.C. (1978).
Representation of complex visual stimuli in the brain.
Naturwissenschaften, 65, 307-318.
The idea was to look for perhaps hidden (non-linear) interactions in complex patterns by systematically scanning
these patterns with the neurons' receptive fields. Responses of many (mainly simple) cells were suprisingly linear,
in correspondence to many other studies since then. The technique provides a nice illustration of the spatial
filtering properties of LGN and cortical cells and of their principle response differences.

•
Nothdurft, H.C., and Lee, B.B. (1982).
Responses to coloured patterns in the macaque lateral geniculate nucleus: Pattern processing in single neurones.
Experimental Brain Research, 48, 43-54.
Study of color representations in LGN cells using Mondrian patterns and the same technique of scanning the
stimuli with the neurons' receptive fields as in the previous work. Provides an illustration of the filter
properties of major cell types in the macaque LGN.
 •
Nothdurft, H.C., and Lee, B.B. (1982).
Responses to coloured patterns in the macaque lateral geniculate nucleus: Analysis of receptive field properties.
Experimental Brain Research, 48, 55-65.
•
Nothdurft, H.C., and Lee, B.B. (1982).
Responses to coloured patterns in the macaque lateral geniculate nucleus: Analysis of receptive field properties.
Experimental Brain Research, 48, 55-65.
(Reverse) reconstruction of the according receptive fields.
 •
Nothdurft, H.C., and Li, C.Y. (1984).
Representation of spatial details in textured patterns by cells of the cat striate cortex.
Experimental Brain Research, 57, 9-21.
•
Nothdurft, H.C., and Li, C.Y. (1984).
Representation of spatial details in textured patterns by cells of the cat striate cortex.
Experimental Brain Research, 57, 9-21.
The challenging observation that certain textures do, and others don't, perceptually segregate set up a project
to study texture segmentation in cortical cells. This paper stresses the uncertainty of representations, texture
orientation within areas, or texture borders between.
•
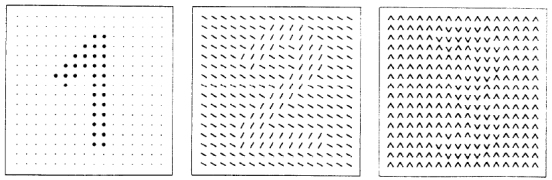
Nothdurft, H.C., and Li, C.Y. (1985).
Texture discrimination: Representation of orientation and luminance differences in cells of the cat striate cortex.
Vision Research, 25, 99-113.
Analysis of texture representation. Both papers together provide nice illustrations of the representation of
orientation differences in cortical cells. They also show, not much stressed at that time, enhanced
(orientation-specific) responses at texture borders—a hint to the later discovered role of feature contrast
in texture segmentation.
Several studies on the spatio-temporal interaction of on and off areas in single neurons were published as conference reports.
2. PSYCHOPHYSICAL (AND PARTLY PHYSIOLOGICAL) STUDIES ON TEXTURE SEGMENTATION:
ANALYSIS OF PROPOSED MODELS
•
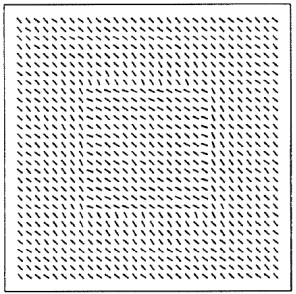
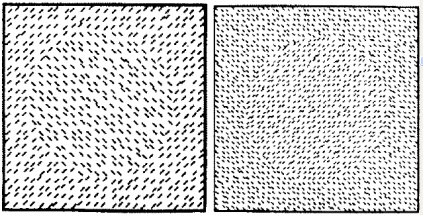
Nothdurft, H.C. (1985).
Sensitivity for structure gradient in texture discrimination tasks.
Vision Research, 25, 1957-1968.
My first strong evidence that texture segmentation is not related to texture features but to gradients of feature
differences. Psychophysics.
•
Nothdurft, H.C. (1985).
Orientation sensitivity and texture segmentation in patterns with different line orientation.
Vision Research, 25, 551-560.
A confirmation of this new view: Texture segmentation from orientation differences is not identical with the
orientation sensitivity, measured at various retinal eccentricities. Psychophysics.
Following the texton model proposed by Bela Julesz, I set up several studies to explore, and eventually demonstrate that the successful segmentation of apparent textons is, in fact, achieved from other stimulus properties. This is addressed in the following three studies.
•
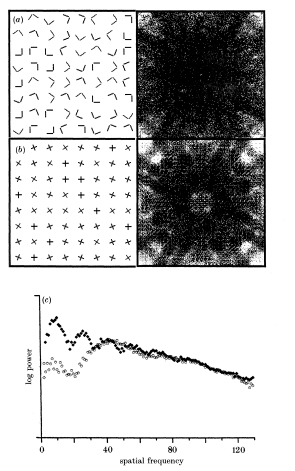
Nothdurft, H.C. (1990).
Texton segregation by associated differences in global and local luminance distribution.
Proceedings of the Royal Society London, B 239, 295-320, and B 241, 249-250.
Shows that "crossings" (a presumed texton property) and "non-crossings" differ in spatial
frequency composition so that variations in spatial configuration that do not affect the distribution of textons
may neverthess render segmentation difficult. Similar demos on other supposed textons. Psychophysics.
 •
Nothdurft, H.C. (1990).
Texture discrimination by cells in the cat lateral geniculate nucleus.
Experimental Brain Research, 82, 48-66.
•
Nothdurft, H.C. (1990).
Texture discrimination by cells in the cat lateral geniculate nucleus.
Experimental Brain Research, 82, 48-66.
Confirmation that many segmented textures are already distinguished at the LGN where neurons are not supposed to
represent the presumed texton filters. Patterns were scanned with the neurons' receptive fields so that response
patterns directly show their responses to various texture areas. Physiology.

•
Nothdurft, H.C. (1991).
Different effects from spatial frequency masking in texture segregation and texton detection tasks.
Vision Research, 31, 299-320.
Another confirmation of the proposal that perceived segmentation is not related to supposed texton differences,
using frequency-band masking. Includes a demo that certain textons can be identified but do not segregate, or
vice versa, with different masking bands. Psychophysics.
3. VARIOUS POPOUT AND SEGMENTATION STUDIES ON SELECTED ASPECTS OF CORTICAL PROCESSING
(PSYCHOPHYSICS)
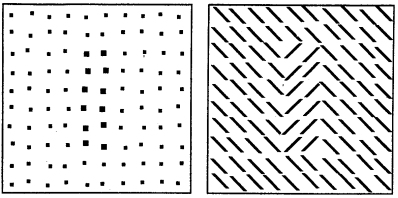 •
Nothdurft, H.C. (1985).
Texture discrimination does not occur at the cyclopean retina.
Perception, 14, 527-537.
•
Nothdurft, H.C. (1985).
Texture discrimination does not occur at the cyclopean retina.
Perception, 14, 527-537.
Following the models of texture processing, it appeared interesting (to me) whether orientation differences
(obviously not represented before the primary visual cortex) do also segregate at the cyclopean retina (which is
also not represented before the primary visual cortex). No, they don't. I found the results quite clear, but
others have later reported evidence for (weak) texture segregation also in stereopatterns.

•
Nothdurft, H.C. (1985).
Discrimination of higher-order textures.
Perception, 14, 539-53.
Illustrates 2nd-order textures of lines and > < items that themselves are only defined by orientation
differences. But careful: Even a small astigmatism will create 1st-order luminance differences between
items and background.
•
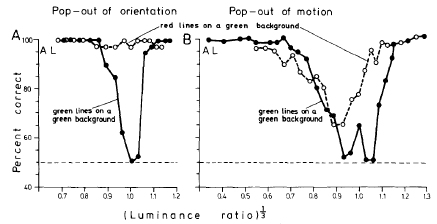
Lüschow, A., and Nothdurft, H.C. (1993).
Pop-out of orientation but no pop-out of motion at isoluminance.
Vision Research, 33, 91-104.
Studies contributions of color sensitive (parvocellular) and color insensitive (magnocellular) processing streams
in different popout tasks.
•
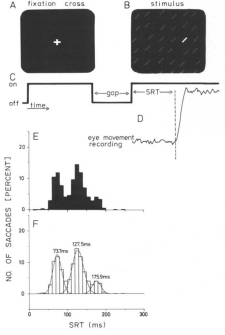
Nothdurft, H.C., and Parlitz, D. (1993).
Absence of express saccades to texture or motion defined targets.
Vision Research, 33, 1367-1383.
The paper addresses a very special phenomenon in vision, the production of "express saccades" under
certain conditions, and studies whether such express saccades may also occur when stimuli are cortically defined
(like motion borders or certain texture borders). We could not find any express saccades to "cortical"
stimuli.

•
Nothdurft, H.C. 1993.
Faces and facial expressions do not pop out.
Perception, 22, 1287-1298.
Given that faces and facial expressions are socially important and are apparently even processed in specialized
cortical areas, this study asked whether or not they do pop out. In my experiments they did not and the detection
of normal faces and facial expressions was not any better than that of scrambled faces or faces presented
upside-down. The study set off a still on-going controverse discussion in the literature whether or not facial
expressions are detected pre-attentively.
4. CONFIRMATION OF THE ORIENTATION CONTRAST MODEL IN TEXTURE SEGMENTATION
(PSYCHOPHYSICS AND PHYSIOLOGY)
 •
Nothdurft, H.C. (1991).
Texture segmentation and pop-out from orientation contrast.
Vision Research, 31, 1073-1078.
•
Nothdurft, H.C. (1991).
Texture segmentation and pop-out from orientation contrast.
Vision Research, 31, 1073-1078.
Shows that popout and segmentation are related to orientation contrast, not orientation per se. Introduces a
paradoxon in which orientation would lead to a different percept than orientation contrast. Psychophysics.
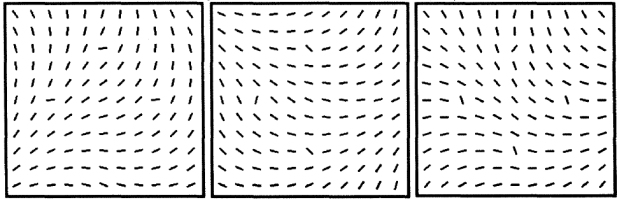
•
Nothdurft, H.C. (1992).
Feature analysis and the role of similarity in pre-attentive vision.
Perception and Psychophysics, 52, 355-375.
More detailed analysis of the previously reported findings, now also including visual search and similarity
grouping. Search of targets with increased orientation contrast is fast, search of targets with similar
orientation contrast as other items is slow. Fast grouping is obtained from (similar) orientation contrast, but
not from similar orientation. The study also addressed alignment effects with oriented lines. Psychophysics.
•
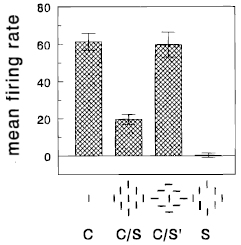
Nothdurft, H.C., Gallant, J.L., and Van Essen, D.C. (1999).
Response modulation by texture surround in primate area V1: Correlates of "popout" under anesthesia.
Visual Neuroscience, 16, 15-34.
Continuation and modification of an earlier study (Knierim & Van Essen, 1992).
The goal was to study the contextual modulation of single cell responses to stimuli within the classical receptive
field by similar or contrasting stimuli in the surrounding regions. Cells were distinguished for their different
type of interactions. Physiology.
•
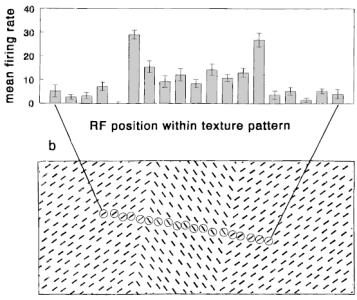
Nothdurft, H.C., Gallant, J.L., and Van Essen, D.C. (2000).
Response profiles to texture border patterns in area V1.
Visual Neuroscience, 17, 421-436.
In-detail analysis of response modulation (by surrounding orientation contrast) of line elements near an
orientation texture border. Observations support the orientation contrast model. Physiology.
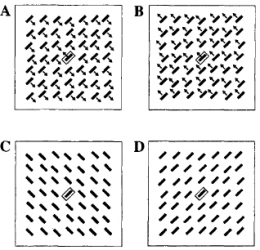
•
Kastner, S., Nothdurft, H.C., and Pigarev, I.N. (1997).
Neuronal correlates of pop-out in cat striate cortex.
Vision Research, 37, 371-376.
Partial replication and extension of the above studies, now investigating orientation and motion popout
from interaction with the surround. Physiology.
•
Kastner, S., Nothdurft, H.C., and Pigarev, I.N. (1999).
Neuronal responses to orientation and motion contrast in cat striate cortex.
Visual Neuroscience, 16, 587-600.
Thorough study of contextual modulation by orientation and motion contrast in the primary visual cortex.
Physiology.
5. TOWARDS A GENERALIZATION OF THE ROLE OF FEATURE CONTRAST (MAINLY PSYCHOPHYSICS)
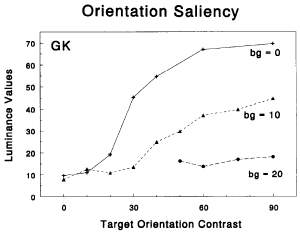
•
Nothdurft, H.C. (1993).
The conspicuousness of orientation and motion contrast.
Spatial Vision, 7, 341-363.
In an attempt to measure the popout strength (salience) of contrasting targets on similar or variable backgrounds,
I matched the salience of an orientation or motion target with that of a luminance target. It turned out that
target salience does not linearly increase with target contrast, and that targets on variable background contrast
(i.e. among distractors that themselves are salient) gradually loose their salience with increasing background
variations.
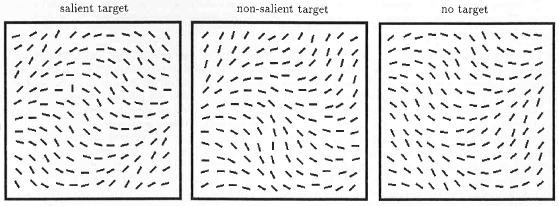
•
Nothdurft, H.C. (1993).
The role of features in preattentive vision: Comparison of orientation, motion, and color cues.
Vision Research, 33, 1937-1958.
The study expands the role of feature contrast beyond orientation, here in motion and color. In all dimensions,
feature contrast lets targets pop out and regions segregate. In grouping, motion behaves like orientation:
Targets with similar (increased) feature contrast are grouped, not targets with similar properties. In color,
however, partial grouping was also obtained from the color itself, not only from color contrast.
(Meanwhile I am uncertain, however, if this was not perhaps due to the usage of certain colors.)
 •
Nothdurft, H.C. (2015).
Feature contrast in salience and grouping: luminance and disparity.
VPL-reports, 3, 132.
•
Nothdurft, H.C. (2015).
Feature contrast in salience and grouping: luminance and disparity.
VPL-reports, 3, 132.
Free download at www.vpl-reports.de/3/
Experiments on two missing feature domains (luminance and disparity) are presented here. The results were similar
to those of the previous study: Feature contrast made targets pop out but targets on varying backgrounds did pop
out less strongly than targets on homogeneous backgrounds. Grouping was seen from the similar salience (feature
contrast) not the similar features of targets, but equal salience was sometimes difficult to achieve. An
additional experiment measured grouping performance with shape-from-shadow figures.
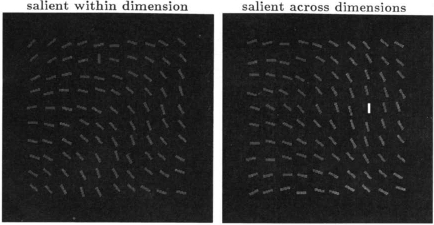
•
Nothdurft, H.C. (1993).
Saliency effects accross dimensions in visual search.
Vision Research, 33, 839-844.
Is the salience effect from orientation contrast specific in visual search? No. It doesn't matter why an oriented
line target is salient. If it is salient, it is found fast, independent of the dimension in which the salience is
generated (by feature contrast). If the target is not salient, search performance is slow.
 •
Nothdurft, H.C. (1994).
Common properties of visual segmentation.
In: Bock, G.R., and Goodie, J.A. (Eds.)
Higher-order processing in the visual system (Ciba Foundation Symposium 184),
Wiley, Chichester, pp. 245-268.
•
Nothdurft, H.C. (1994).
Common properties of visual segmentation.
In: Bock, G.R., and Goodie, J.A. (Eds.)
Higher-order processing in the visual system (Ciba Foundation Symposium 184),
Wiley, Chichester, pp. 245-268.
The article summarizes the current findings from my lab and gives several demos on the functional similarity of orientation
and luminance contrast in segmentation.
•
Nothdurft, H.C. (1994).
Cortical properties of preattentive vision.
In: Albowitz, B., Albus, K., Kuhnt, U., Nothdurft, H.C., and Wahle, P. (Eds.)
Structural and functional organization of the neocortex,
Springer Verlag Heidelberg, pp. 375-384.
The study summarizes the phenomena of popout, segmentation, and grouping with differently oriented lines.
•
Nothdurft, H.C. (1997).
Different approaches to the encoding of visual segmentation.
In: Harris, L., and Jenkins, M. (Eds.)
Computational and psychophysical mechanisms of visual coding,
Cambridge University Press, New York, pp. 20-43.
Another review of segmentation studies from my lab. The paper summarizes the major properties and provides demos
of segmentation effects that should not be expected from other models. Last not least, the paper presents results
from a computational model based on contextual, feature-specific suppression (as found in the physiological
studies above) that does predict several observed perceptual phenomena.
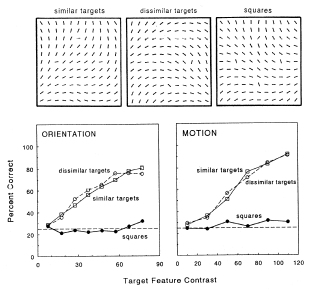
The following three papers represent the data from a larger psychophysical project in which salience from feature contrast was studied, and compared, between various dimensions: orientation, luminance, color, and the direction of motion. The major findings were summarized for a textbook article.

•
Nothdurft, H.C. (2000).
Salience from feature contrast: additivity across dimensions.
Vision Research, 40, 1183-1201.
Do salience effects from different dimensions, e.g. orientation and luminance, add? Yes, they do, but non-linearly
and not with the same magnitudes for different features (as one should expect from the properties of
cortical neurons).
 •
Nothdurft, H.C. (2000).
Salience from feature contrast: temporal properties of saliency mechanisms.
Vision Research, 40, 2421-2435.
•
Nothdurft, H.C. (2000).
Salience from feature contrast: temporal properties of saliency mechanisms.
Vision Research, 40, 2421-2435.
Can we distinguish the timings of feature contrast mechanisms in different feature domains? Yes, we can. This was
studied by measuring the temporal resolution of alternating segregation patterns. The segmentation of luminance
and color differences could be followed faster than that of orientation and motion differences.
 •
Nothdurft, H.C. (2000).
Salience from feature contrast: variations with texture density.
Vision Research, 40, 3181-3200.
•
Nothdurft, H.C. (2000).
Salience from feature contrast: variations with texture density.
Vision Research, 40, 3181-3200.
Does the salience of target contrast depend on the spatial configuration of items? Yes, it does. Dense (but not
too dense) configurations of feature differences do generally produce stronger salience than sparse configurations.

•
Nothdurft, H.C. (2005).
Salience of feature contrast.
In: Itti, L., Rees, G., and Tsotsos, J. K. (Eds.)
Neurobiology of attention, Elsevier, San Diego, CA, pp. 233-239.
This textbook article illustrates various examples of target salience and summarizes findings of the three
articles before.
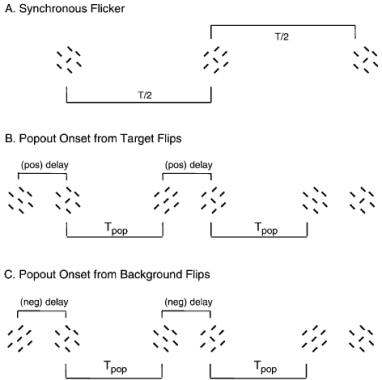
•
Nothdurft, H.C. (2002).
Latency effects in orientation popout.
Vision Research, 42, 2259-2277.
The different timing of central responses and contextual suppression in cortical neurons (as seen, for example,
in the physiology studies above) predicts perceptual differences depending on whether the (oriented) target or
the orthogonal surround precede in presentation. The study investigates timing effects in two ways. It first
looks for principle differences between the detection and discrimination of single lines and oriented line
texture fields. Secondly it measures the detectability of asynchronously presented targets and surrounds and
compares target detection rates with predictions from a model with different delays for responses and contextual
suppression.
6. ATTENTION SHIFTS IN VISUAL SEARCH (PSYCHOPHYSICS)

•
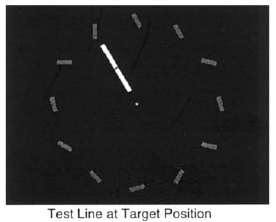
Ziebell, O., and Nothdurft, H.C. (1999).
Cueing and pop-out.
Vision Research, 39, 2113-2125.
Study on the dynamics of popout from orientation and on the interaction of cues and salience.
•
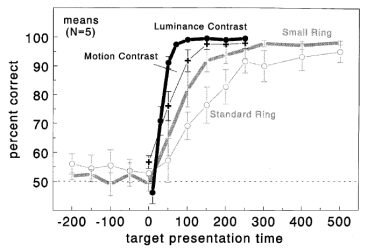
Nothdurft, H.C. (1999).
Focal attention in visual search.
Vision Research, 39, 2305-2310.
An open question in earlier models was whether the so-called pre-attentive visual search was indeed free of
attention, and whether presumably serial search performance was indeed associated with shifts of attention. This
study measured attention shifts to targets and found that even in "pre-attentive" search the target
was attended to.
•
Nothdurft, H.C. (2002).
Attention shifts to salient targets.
Vision Research, 42, 1287-1306.
Presumed shifts of attention were studied with various targets and saliency markers. The results underline the
general role of salience in controlling and guiding attention within a pattern.

•
Nothdurft, H.C. (2006).
Salience and target selection in visual search.
Visual Cognition, 14(4-8), 514-542.
This special role of salience is here stressed and further analyzed. Salience is one of the key features in
visual search. The paper shows that the detection of salient items is faster than their identification, and that
the search time needed to find a target in large assemblies is controled by the number of salient elements.
Additional papers on salience-controlled attention shifts ("visual selection") are in preparation.
7. TOWARDS STUDYING ATTENTION SHIFTS IN VISUAL SEARCH (PHYSIOLOGY)
In an earlier research project at the Max-Planck-Institute for Biophysical Chemistry in Göttingen, attention shifts were also studied in single cells. The following papers of the project were published.
 •
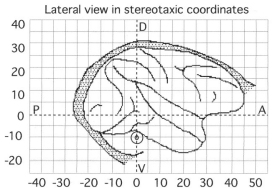
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (1997).
Evidence for aynchronous development of sleep in cortical areas.
Neuroreport, 8, 2557-2560.
•
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (1997).
Evidence for aynchronous development of sleep in cortical areas.
Neuroreport, 8, 2557-2560.
An observation of "local" sleep during the performance of a highly trained visual task.
•
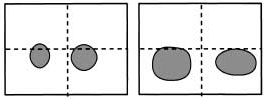
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (1997).
A reversible system for chronic recordings in macaque
monkeys.
Journal of Neuroscience Methods, 77, 157-162.
Development of a new and reversible technique to implant a holder for head stabilization and recordings in
intermediate chronic experiments.
 •
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (2001).
Neurons with large bilateral receptive fields in
monkey
prelunate gyrus.
Experimental Brain Research, 136, 108-113.
•
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (2001).
Neurons with large bilateral receptive fields in
monkey
prelunate gyrus.
Experimental Brain Research, 136, 108-113.
Report of neurons with large receptive fields reaching across the vertical meridian.
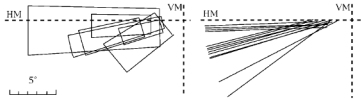 •
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (2002).
Neurons with radial receptive fields in
monkey
area V4A: evidence of a subdivision of prelunate gyrus based on neuronal response properties.
Experimental Brain Research, 145, 199-206.
•
Pigarev, I.N., Nothdurft, H.C., and Kastner, S. (2002).
Neurons with radial receptive fields in
monkey
area V4A: evidence of a subdivision of prelunate gyrus based on neuronal response properties.
Experimental Brain Research, 145, 199-206.
Report of a group of neurons with particular response properties.
•
Nothdurft, H.C., Pigarev, I.N., and Kastner, S. (2009).
Overt and covert visual search in primates: Reaction times and gaze shift strategies.
Journal of Integrative Neuroscience, 8(2), 137174.
Overview of various behavioral observations in overt and covert search tasks.
8. LUMINANCE-DEFINED SALIENCE (PSYCHOPHYSICS)
•
Nothdurft, H.C. (2006).
Salience-controlled visual search: are the brightest and the least bright targets found by different processes?
Visual Cognition, 13(6), 700-732.
Following the previous conclusions on the important role of salience in visual search, a peculiar
assumption from the literature was questioned. The brightest and the dimmest targets (in an esemble of bright
targets) were assumed to be searched for in different processes. The brightest target (among less bright targets)
was believed to be found in parallel search, while the least bright target (among brighter targets) was believed
to be found in serial search. The present experiments showed that these two targets differ in salience and that,
when saliences are adjusted, both targets are found in a similar (seemingly "parallel") search process.
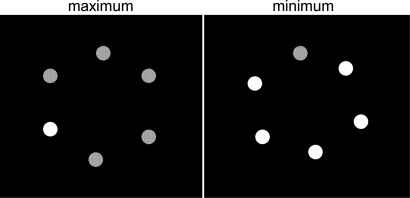
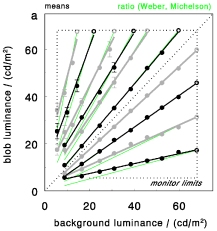
These observations had led to study the properties of luminance-defined salience in more detail, in the following two papers. In two blob arrays, targets were matched for salience, and resulting luminance settings were analyzed. Unfortunately, the findings were less simple and less clear-cut than hoped, but they still provided several principle rules.
•
Nothdurft, H.C. (2015).
Luminance-defined salience of homogeneous blob arrays.
VPL-reports, 1, 138.
Free download at www.vpl-reports.de/1/
The salience of single targets or arrays of similar items follows the Weber contrast, unless targets are (much)
brighter than background. When backgrounds differ considerably, matches of bright and dark targets may follow
Steven's brightness law instead.
 •
Nothdurft, H.C. (2015).
Luminance-defined salience - targets among distractors.
VPL-reports, 2, 197.
•
Nothdurft, H.C. (2015).
Luminance-defined salience - targets among distractors.
VPL-reports, 2, 197.
Free download at www.vpl-reports.de/2/
In matches of single targets among (different) distractors, various cases must be distinguished. Dark targets
tend to be similarly salient, if their luminance difference to distractors is identical (irrespective of the
background). Bright targets often appear similarly salient, if their luminance difference to distractors
(eventually normalized to the luminance span of background and distractors) is the same. But very bright targets
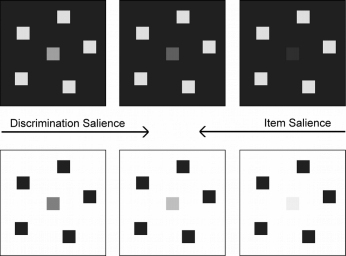
may behave differently. Matches of bright and dark targets are strongly controlled by Steven's law. The paper
also distinguishes between item salience (contrast to background) and discrimination salience (contrast between
targets and distractors).
9. CUED VISUAL SELECTION (PSYCHOPHYSICS)
Several studies developped from the above paper on Attention shifts to salient targets (2002). The first two publications are the earliest follow-ups (from 2003-2004) which were, however, not published before 2016. More publications are "in preparation".
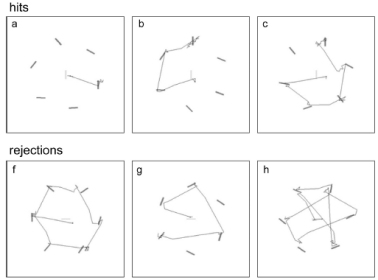
•
Nothdurft, H.C. (2016).
Spatial cue effects in Visual Selection
VPL-reports, 4, 121.
Free download at www.vpl-reports.de/4/
 This paper reports measurements of spatial cue effects. Targets were marked by a cue. Near cues interacted
with target perception independent of the selection process. Far cues delayed the selection process.
In addition, the effects of figural connections upon the cue-target linking were investigated ("tentacles").
This paper reports measurements of spatial cue effects. Targets were marked by a cue. Near cues interacted
with target perception independent of the selection process. Far cues delayed the selection process.
In addition, the effects of figural connections upon the cue-target linking were investigated ("tentacles").

•
Nothdurft, H.C. (2016).
Fast identification of salient objects depends on cue locations.
VPL-reports, 5, 18.
Free download at www.vpl-reports.de/5/
The efficiency of target identification (here, line orientation) inside and outside selected objects (here, boxes)
was measured as a function of the marker location upon the boxes' frame.
•
Nothdurft, H.C. (2017).
Cued visual selection a tool to study the dynamics of neural processes in perception?
VPL-reports, 6, 124.
Free download at www.vpl-reports.de/6/
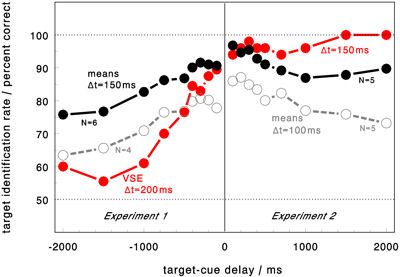 The paper studies the dynamics of Cued Visual Selection (CVS). Beside the standard cuing paradigm (cues first,
then targets) also the reversed sequence was tested, in which cues were later superimposed upon the test pattern.
Target identification underlies interesting variations aith increasing delays. The paper gives an introduction,
particularly to the latter variation of the paradigm, which appears to be a useful tool to look into the dynamics
of neural processes with psychophysical tools.
The paper studies the dynamics of Cued Visual Selection (CVS). Beside the standard cuing paradigm (cues first,
then targets) also the reversed sequence was tested, in which cues were later superimposed upon the test pattern.
Target identification underlies interesting variations aith increasing delays. The paper gives an introduction,
particularly to the latter variation of the paradigm, which appears to be a useful tool to look into the dynamics
of neural processes with psychophysical tools.
•
Nothdurft, H.C. (2017).
Cued visual selection of targets with and without orientation contrast.
VPL-reports, 7, 122.
Free download at www.vpl-reports.de/7/
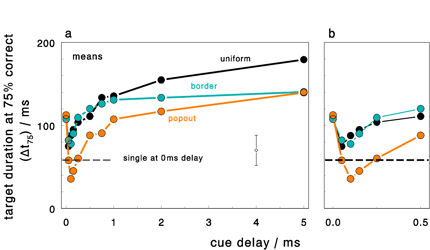
The efficiency of target identification (oriented lines) was tested with CVS at various cue delays after stimulus
onset. As predicted from neural population responses, the local target context has an important influence.
Targets in popout configurations are faster identified than targets in uniform or border configurations.
•
Nothdurft, H.C. (2018).
Cued visual selection in binocular rivalry.
VPL-reports, 8, 132.
Free download at www.vpl-reports.de/8/
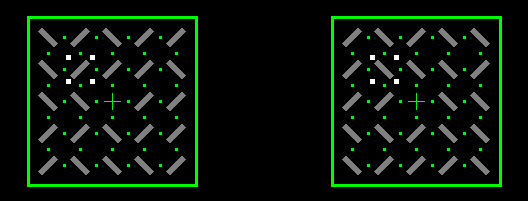 In line rasters with orthogonal orientations in the two eyes, the detection of monocularly and binocularly cued
targets was measured and by and large reflects the activity variations expected for the changing ocular patterns.
However, except for the first 500-1000ms after pattern onset, when ocular interactions
seem to develop and settle, modulations are generally too small to explain the known alternating and exclusive
percepts in binocular rivalry.
In line rasters with orthogonal orientations in the two eyes, the detection of monocularly and binocularly cued
targets was measured and by and large reflects the activity variations expected for the changing ocular patterns.
However, except for the first 500-1000ms after pattern onset, when ocular interactions
seem to develop and settle, modulations are generally too small to explain the known alternating and exclusive
percepts in binocular rivalry.
•
Nothdurft, H.C. (2018).
Time window(s) of cued visual selection.
VPL-reports, 9, 131.
Free download at www.vpl-reports.de/9/
The time course of cued target identification was measured in different feature dimensions.
Cued targets were generally identified in short time windows which varied between features and observers, but
attention could also be held for later targets if the cued location was empty or the requested information
not yet available.
For orientation, color, and luminance polarity, but not for motion, target properties were faster seen than
cues detected; instead of reporting the properties of the physically cued target observers reported the
properties of a later target at the cued location. That is, the effective cuing windows were delayed.
Different directions of target movement, however, were identified about in synchrony with the cue.
These differences confirm earlier reports of an asynchronous perception of color, orientation, and motion.
•
Nothdurft, H.C. (2019).
Location-cued visual selection placeholder dots improve target identification.
Journal of Vision, 19(13):16, 115, https://doi.org/10.1167/19.13.16
Free download at
https://jov.arvojournals.org/article.aspx?articleid=2755884

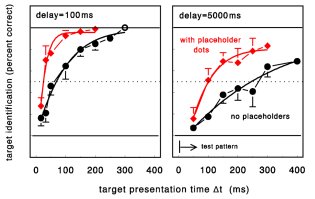 Briefly presented cues generate a transient benefit in target identification (delays 50300ms)
followed by a continuous decay of performance when targets were onset later (measured up to
a delay of 5s). The decay is substantially reduced by placeholder dots shown during the delay.
Analysis of target presentation times needed for constant performance (e.g., 75%) reveals the strengths
of the underlying (neural) signals. The effect of placeholder dots is then seen as a nearly constant
addition to the cued target signals, with an additional transient peak about 100 ms after cue and
placeholders onset.
Briefly presented cues generate a transient benefit in target identification (delays 50300ms)
followed by a continuous decay of performance when targets were onset later (measured up to
a delay of 5s). The decay is substantially reduced by placeholder dots shown during the delay.
Analysis of target presentation times needed for constant performance (e.g., 75%) reveals the strengths
of the underlying (neural) signals. The effect of placeholder dots is then seen as a nearly constant
addition to the cued target signals, with an additional transient peak about 100 ms after cue and
placeholders onset.
 •
Nothdurft, H.C. (2020).
Dynamic differences in the identification of orientation, color, and facial expressions.
VPL-reports, 10, 127.
•
Nothdurft, H.C. (2020).
Dynamic differences in the identification of orientation, color, and facial expressions.
VPL-reports, 10, 127.
Free download at www.vpl-reports.de/10/
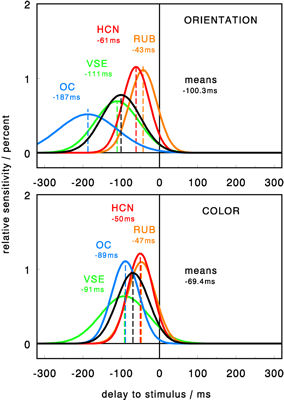
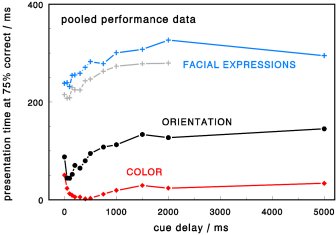
The time course of cued target identification was measured for orientation, color, and facial
expressions. Color was identified first, followed by orientation, then facial expressions.
Performance variations give insights into the dynamics of underlying neural signals. Color is encoded
fast, with a peak still increasing up to 400 ms and a strong sustained response component. Orientation
is also encoded fast, with an earlier peak at 50-100 ms and a primarily transient response. Signals
representing facial expressions were generally weaker (and delayed) but seem to share the dynamic
shape of orientation channels.
•
Nothdurft, H.C. (2020).
Cued visual selection of conjunction targets no evidence of additional attentional requirements for the binding of color and orientation.
VPL-reports, 11, 114.
Free download at www.vpl-reports.de/11/
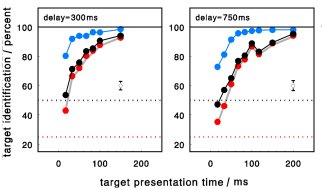 When red and green lines have to be discriminated, their color is faster identified than their
orientation. Thus, even in conjunction tasks, features are processed separately and independently.
Too short target presentations may produce "false" conjunctions, but not because of a lack of
attentive feature integration but merely because of too slow feature identification processes.
Nevertheless, the identification of target orientation may be faster in conjunction than in
single-feature tasks, suggesting that there is no need of additional attentional resources
for feature binding on top of the attentional capacities required for target identification.
When red and green lines have to be discriminated, their color is faster identified than their
orientation. Thus, even in conjunction tasks, features are processed separately and independently.
Too short target presentations may produce "false" conjunctions, but not because of a lack of
attentive feature integration but merely because of too slow feature identification processes.
Nevertheless, the identification of target orientation may be faster in conjunction than in
single-feature tasks, suggesting that there is no need of additional attentional resources
for feature binding on top of the attentional capacities required for target identification.
•
Nothdurft, H.C. (2020).
Distance versus hemifield costs in the identification of cued double targets.
VPL-reports, 12, 132.
Free download at www.vpl-reports.de/12/
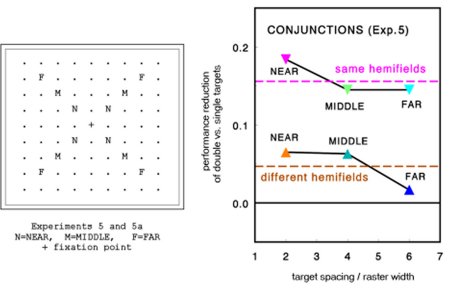 The paradigm of Cued Visual Selection was used to measure the identification speed of single
targets and target pairs in large item arrays. Four target categories were tested: oblique lines,
Vernier's, T letters at different orientations, and conjunctions of orientation & color.
All targets were briefly cued. Target pairs in different distances from another and located in same
or different visual hemifields were compared with the identification rates of single targets at same
eccentricities. There was a clear predominance of hemifield effects: The identification of target
pairs in same hemifields was notably worse than the identification of target pairs in different
hemifields.
The paradigm of Cued Visual Selection was used to measure the identification speed of single
targets and target pairs in large item arrays. Four target categories were tested: oblique lines,
Vernier's, T letters at different orientations, and conjunctions of orientation & color.
All targets were briefly cued. Target pairs in different distances from another and located in same
or different visual hemifields were compared with the identification rates of single targets at same
eccentricities. There was a clear predominance of hemifield effects: The identification of target
pairs in same hemifields was notably worse than the identification of target pairs in different
hemifields.
10. MORE ABOUT TEXTURE: TEXTURE IDENTIFICATION
 •
Nothdurft, H.C. (2024).
Low sensitivity for orientation in texture similarity ratings.
Journal of Vision, 24 (13):14, 1-9. doi:https://doi.org/10.1167/jov.24.13.14.
Open Access: https://jov.arvojournals.org/article.aspx?articleid=2802383
•
Nothdurft, H.C. (2024).
Low sensitivity for orientation in texture similarity ratings.
Journal of Vision, 24 (13):14, 1-9. doi:https://doi.org/10.1167/jov.24.13.14.
Open Access: https://jov.arvojournals.org/article.aspx?articleid=2802383
Early report of two experiments on texture identification, in which the perceived ranking of
features was studied with similarity ratings of texture fields. A particularly low ranking
was found for orientation, although orientation variations themselves are easily segregated.
Two test series are reported here, a combination of color and line orientation, and a combination
of spatial frequency and texture orientation. In both experiments, observers tended to ignore
orientation when grouping texture patches for similarity, and preferred similarities in the second
dimension, color or spatial frequency, even across different orientations.
 •
Nothdurft, H.C. (2025).
Perceived ranking of feature dimensions in texture similarity ratings.
VPL-reports, 13, 1-19. doi: 10.4126/FRL01-006488495 (via repository.publisso.de)
Free download at www.vpl-reports.de/13/
•
Nothdurft, H.C. (2025).
Perceived ranking of feature dimensions in texture similarity ratings.
VPL-reports, 13, 1-19. doi: 10.4126/FRL01-006488495 (via repository.publisso.de)
Free download at www.vpl-reports.de/13/
Combinations of plain or textured patterns were presented to human observers to see which features
are most important for texture identification and which ones might eventually be ignored. Pairwise
ranking of features and feature domains was measured
from spontaneous decisions on the similarity of simultaneously displayed test patches.
 In plain patches differing in color or luminance, color
predominated the similarity ratings until luminance or saturation differences became very strong and affected the
perceived hue of color patches. In line textures, both color and luminance were more important than
any of the spatial features line size, line density, and line orientation. In textures without color
or luminance variations, line size and density were more important than orientation. Altogether, the
results reveal a systematic ranking of features in similarity estimates that was, with gradual
variations, seen across all observers. Color was the most important and, surprisingly, orientation
the least important feature domain.
In plain patches differing in color or luminance, color
predominated the similarity ratings until luminance or saturation differences became very strong and affected the
perceived hue of color patches. In line textures, both color and luminance were more important than
any of the spatial features line size, line density, and line orientation. In textures without color
or luminance variations, line size and density were more important than orientation. Altogether, the
results reveal a systematic ranking of features in similarity estimates that was, with gradual
variations, seen across all observers. Color was the most important and, surprisingly, orientation
the least important feature domain.